
CSEC Biology: Reproduction in Plants
- JH@Quelpr

- Apr 22, 2020
- 6 min read
Updated: Apr 23, 2020
Plant reproduction is far different from reproduction in any animal. First of all, plants use flowers as their reproductive organs (which is sort of strange if you think about it), and their fertilization is known as pollination. You should also note that not all plants are restricted to sexual reproduction using flowers and seeds, and some actually use asexual reproduction which we discuss in this other lesson.
As we said before, the main site of plant reproduction is in the flower. Each flower has small differences based on what suits the plant best, however most flowers have 10 basic parts (which CSEC requires you to know). These 10 parts are best split up into 4 circular sections or whorls: the calyx, corolla, stamen (or androecia) and pistil (or gynoecia).
The parts of the flower are as follows:
Sepals- The sepals are the green-coloured part towards the base of the flower connected to the receptacle that holds together the petals and protects the developing bud of the flower from mechanical damage and drying out (dessication). The receptacle mentioned is the thickened part of a stem from which the flower organs grow. The sepals are considered to be the calyx of the flower, which is the outermost whorl.
Petals- The petals are the 2nd whorl of the flower, or the corolla. The petals are the most visible part of the flower, and are usually brightly coloured and have fragrances to attract pollinators such as birds, bees and insects (please also note that not all flowers are insect-pollinated, and we will discuss the difference in structure later on). The petals also provide protection to the reproductive inner part of the flower. Not only this, but the petals work as a sort of platform on which insects can land. The calyx and corolla are collectively called the perianth.
The third whorl of the flower, the stamen, is the male portion of the flower, and consists of two main parts:
Anther- The anther produces pollen grains, which contain the male gametes of the flower necessary for sexual reproduction. The anther also stores these pollen grains until they are ready for dispersal. One flower's stamen can contain several groups of anthers.
Filament- The filament is a stalk-like structure which attaches to the base of the flower (receptacle) and supports the anther.
The fourth whorl of the flower, the pistil, is the female reproductive portion of the flower,
Stigma- This is a sticky tissue at the end of the pistil that is receptive to pollen during pollination.
Style- The style is a long rod-shaped tube that extends from the ovary in the base of the flower to the stigma higher up in the flower. The style usually supports the stigma to a height above other components of the flower so that the stigma can catch pollen more easily.
Ovary- As previously mentioned, the hollow tube of the style leads into the ovary. The ovary is the swollen base portion of the pistil, which functions as a protective vessel for the ovules within it. When pollen enters the ovary through the style and therefore fertilizes the ovules, the ovary develops into a fruit. This fruit protects the developing seeds to maturity until the fruit itself becomes a vehicle for the dispersal of the seeds (mature ovules).
Ovules- The ovules are the sites at which the female reproductive cells are produced (and contained). After fertilization, the ovules develop into seeds, which can eventually germinate and grow into a new plant.
Embryo sac- The embryo sac, also known as the female gametophyte, is where the egg cell of the plant will be fertilized and the embryo will develop. It consists of 6 haploid cells without cell walls.
Micropyle- The micropyle has an additional function in germination, but in the reproductive aspect of the plant, the micropyle is the small opening in the ovule through which pollen enters during pollination.

Flowers can either be insect-pollinated or wind-pollinated. In insect pollination, an insect will carry the pollen from one flower to another. However, in wind pollination, the pollen from the anthers is instead picked up in wind currents and caught by other flowers' stigmas. Due to the different methods of pollination, these flowers have structures and adaptations suited best to each.
Insect Pollinated Plants Insect pollinated plants have flowers structurally adapted to attract insects. So, these flowers tend to produce sweet nectar as incentive for insects to land on the flower and drink the nectar, picking up its pollen in the process and carrying it around to other flowers. The diagram above closely resembles an insect-pollinated flower.
These flowers will also have large brightly coloured and scented/fragrant petals to attract insects.
The petals themselves may have structures known as nectar guides, which are markings that direct pollinators to the nectar.
The pollen grains of these plants also tend to be large and sticky or spiky to better adhere to the insects.
The anthers will produce a smaller amount of pollen grains since few grains will be lost.
The stigmas of these flowers are usually small, compact and do not protrude out of the flower.
The stigmas are not feathery and are instead sticky, so that the pollen grains are not easily dispersed.
The stamen is located within the interiors of the flower, so that pollen can stick on the backs of pollinators as they brush against the anthers.
Wind Pollinated Plants
Wind pollinated plants do not rely on attracting insects to get pollen around. Instead, their structure is adapted to make it easier for the wind to pick up their pollen and transport it between flowers.
The flowers are usually small, dull-colored and scentless and therefore, not attractive to insects.
Nectar is usually absent, as again, there is no need to attract insects.
Pollen is very abundant as wastage is higher. That is, the flower will produce more pollen since lots of it will be lost in the wind.
Pollen grains are small, dry, smooth and light so that they are buoyant and easily blown about by air currents.
Stamens usually have long, slender filaments that sway in the wind easily, allowing pollen grains to be shaken out from the anthers as a result.
Stigmas protrude from the flower itself and are large and feathery so that they provide a large surface area to catch pollen floating in the air.
Nectar guides are absent

Pollination is the process by which the pollen grains of the male anther of a flower reach the female stigma. This can either occur through self-pollination or cross-pollination.
Self-pollination is when the pollen grains from the anther of a flower reach the stigma of the same flower or another flower on the same plant.
Cross-pollination is when the pollen grains from the anther of a flower reach the stigma of another flower on a different plant of the same species.
In comparison, fertilization occurs in two main steps, pollen grain germination and double fertilization.
First, the pollen grain germinates after pollination and grows into the style by creating the pollen tube (a pathway for the pollen grain to move down to the ovary). The pollen tube breaks into the ovule through the micropyle and bursts into the embryo sac. This is where the fusion of the gametes occurs.
Then, double fertilization occurs- which comprises two stages: the fertilization of the ovule and triple fusion. Double fertilization starts with the fertilization of the ovule, where one of the two male nuclei of the pollen grain fuse with the egg nucleus to form the zygote, which divides over time, forming the embryo. Then comes triple fusion, which is the fusion of the other male nucleus with the two embryo sac nuclei, forming the endosperm nucleus which divides to form the endosperm tissue.
Here is a diagram that might simplify the whole process:

(You can read more here, for a more complex explanation)
CSEC also requires that you understand the process of seed and fruit formation in dicotyledons.
The endosperm starts developing immediately after double fertilization, forming a tissue between the ovule and the young embryo (the endosperm divides actively during early development of the seed but later, it may store considerable amounts of food material in endospermic seeds, or it may transfer the food materials to the cotyledon(s) of the embryo and disappear completely in non-endospermic seeds). Meanwhile, the zygote undergoes a regular sequence of mitotic divisions to form an embryo organized into a central hypocotyl with a radicle at one end and one or two cotyledons (in dicots) and small shoot meristem (plumule) at the other. The outside (integuments) of the ovule become modified to form the seed coat (testa). The entire structure--seed coat, endosperm, and embryo-constitutes the seed.
After this, the seed dries out (dessication). Water is drawn from the seed, causing it to become dormant.
Seed development in dicotyledons (and any plant) is a quite complex process involving histodifferentiation (embryogenesis), cell expansion and maturation drying. We recommend that you look at this explanation of embryogenesis if you want a more in-depth look at the process.
Here's a little fun fact that you probably did not know before:
The tissue that attached the seed to the mother plant (fruit) is called the funiculum. It is
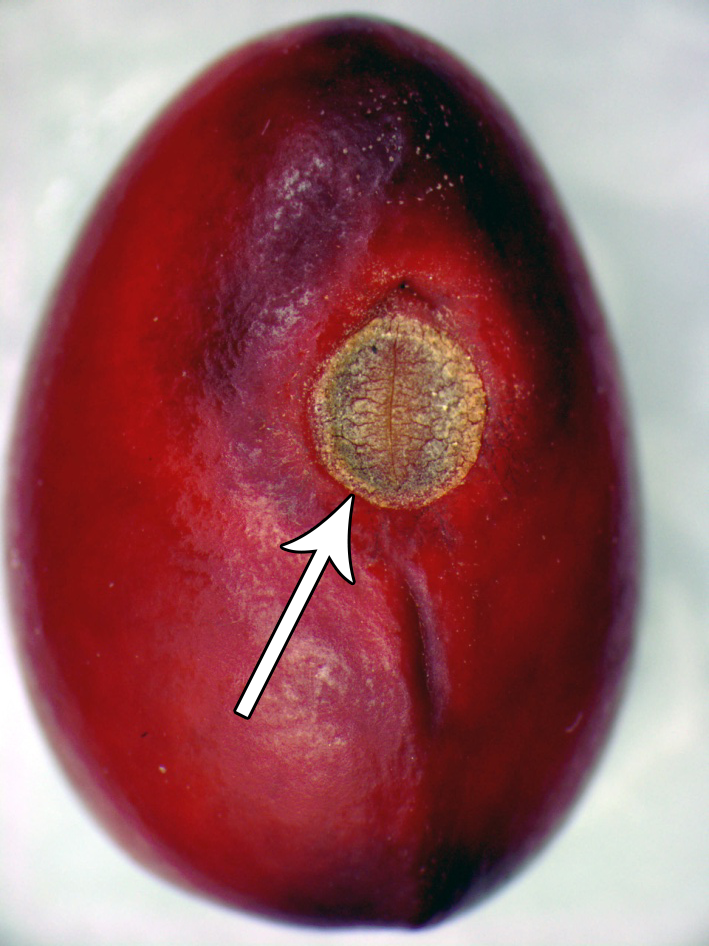
the conduit through which food and water reached the seed. However, right before seed dessication, or mature drying when the seed reaches maturity, the funiculum is severed, leaving a scar known as a hilum.
The walls of the ovary develop into the fruit through mitotic divisions. The stigma, petals, style (and sepals in some fruits) will wilt and fall off. The fruit can either be dehiscent (dry) or indehiscent (succulent) based on the pericarp. The pericarp is the wall of the fruit composed of 3 layers: the exocarp, mesocarp and endocarp.
So, in indehiscent fruits, the pericarp has one or more fleshy or juicy layers and in dehiscent fruits, the pericarp is thin and dry.



Kaiser OTC benefits provide members with discounts on over-the-counter medications, vitamins, and health essentials, promoting better health management and cost-effective wellness solutions.
Obituaries near me help you find recent death notices, providing information about funeral services, memorials, and tributes for loved ones in your area.
is traveluro legit? Many users have had mixed experiences with the platform, so it's important to read reviews and verify deals before booking.